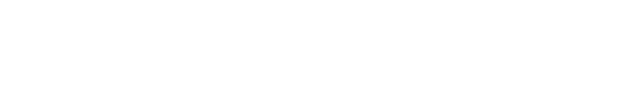

慢性疼痛におけるDRGニューロン遠位末端(皮膚)の変化
DRGニューロン(C線維、偽単極性ニューロン、一次感覚ニューロン)

慢性痛におけるDRGニューロンの遠位末端(皮膚)
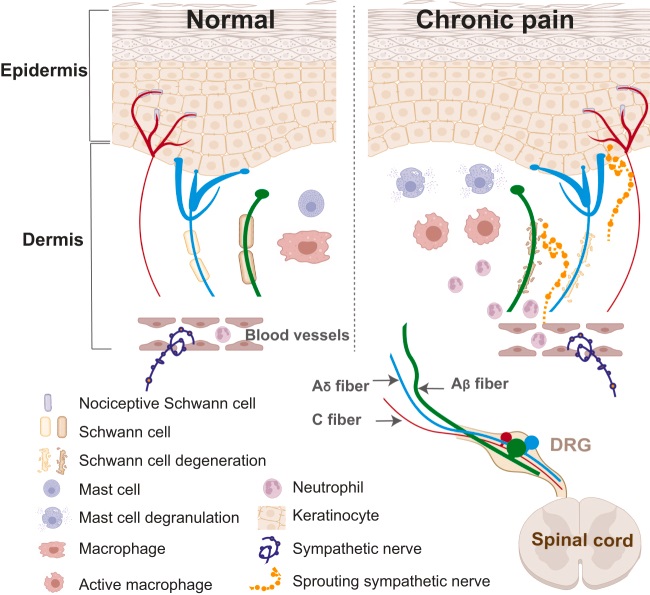
図:痛みに関与する末梢組織の主な細胞タイプ
図の説明:
「痛みの感覚情報は、主に小径線維(AδタイプとCタイプ)の感覚ニューロンにより受け取られる。
感覚ニューロンの末梢軸索は、真皮(Dermis)と表皮(Epidermis)の両方に伸びる精巧な終末枝を形成し、細胞表面レセプタを介して、
多種多様な刺激を受ける。
生理的な条件下では、微小環境はダイナミックバランスにある。
慢性疼痛状態では、角化細胞(ケラチノサイト:Keratinocyte)、肥満細胞(Mast cell)、マクロファージ(Macrophage)が活性化され、
侵害ニューロンを刺激または感作するメディエーターを放出する。
好中球(Neutrophil)のような、循環する免疫細胞は、血流からリクルートされる。
通常、軸索束を包むシュワン細胞は変性する。
さらに、通常は、真皮を支配しない、交感神経が、真皮上部と表皮に芽生え、感覚システムと直接相互作用する。」
上皮細胞の役割:
皮膚のケラチノサイト、腸やその他の内臓の上皮細胞タイプを総称して上皮細胞と呼ぶ。DRGニューロンの末梢末端はケラチノサイトまで伸びている。上皮細胞が傷害されると、痛みの原因となるさまざまな神経活性メディエーターを分泌する。
これらには、アデノシン5'-3リン酸(adenosine 5'-triphosphate: ATP)、神経成長因子(nerve growth factort: NGF)、インターロイキン-6(interleukin-6: IL-6)、インターロイキン-1(interleukin-1: IL-1)、腫瘍壊死因子α(tumor necrosis factor α: TNF-α)、エンドセリン-1(endotrhelin-1: ET-1)が含まれる。
[ATP]
(1)皮膚へのATP投与は、ヒトと動物の両方で疼痛行動を誘発する。
(2)無害もしくは有害な触覚は、ケラチノサイトからのATPを放出し、ケラチノサイトは、感覚ニューロン上の、P2X4イオンチャネル受容体に直接作用する。
(3)帯状疱疹後神経痛や複合性局所疼痛症候群などの疾患では、表皮のATP放出が増加し、感覚ニューロン上の、P2X受容体の過剰な活性化を引き起こし、疼痛を誘発する可能性がある。
(4)ATPは、感覚ニューロン上の、プリン作動性受容体(P2Y)Gタンパク質共役型受容体(G-protein-coupled receptor:GPCR)に作用することができる。
(5)P2Y受容体の根本的な興奮機構は、TRPV1、カリウムチャネル(Kv7)、メカノトランスダクションチャネルなどのイオンチャネルの調節によって誘導される可能性がある。
[NGF]
もともと、発生過程における神経細胞生存因子として発見された。
(1)NGFは侵害受容器ニューロンに発現する高親和性受容体である、トロポミオシン受容体キナーゼA(tropomyosin receptor kinase A: Trk A)に結合する。
(2)NGF/TrkAシグナルは、TRPV1を感作し、Nav1.8 Na+チャネルをコードする遺伝子(SCN10a)、酸感受性イオンチャネル3(acid-sensing ion channel 3: Accn3)、TRPV1受容体、脳由来神経栄養因子(brain derived neurotrophic factor: BDNF)、サブスタンスP(substance P)などの遺伝子を発現させる。
[IL-6]
神経造血系サイトカインファミリーである。
(1)IL-6の足底内注射は、用量依存的に、機械的痛覚過敏を誘発する。
(2)IL-6は、培養DRGニューロンの約33%で、急速なカルシウム過度現象を引き起こすことができるので、疼痛ニューロンに対するIL-6の効果は直接的かもしれない。
(3)IL-6は、可溶性IL-6レセプターとgp130を介して神経細胞を活性化し、Grb2関連結合様タンパク質(Gab)1/2/ホスファチジルイノシトール3キナーゼ(PI3K)を介して、プロテインキナーゼC(PKC)-δを活性化することにより、TRPV1のリン酸化を促進することができる。
(4)IL-6/可溶性IL-6受容体/gp130シグナルは、熱誘発性CGRP放出を増強することができる。
[IL-1b と 腫瘍壊死因子α(tumor necrosis factor α: TNF-α)]
炎症性サイトカインである。
(1)IL-1bまたはTNF-αの足底注射は、機械的痛覚過敏と熱的痛覚過敏の両方を引き起こすことができる。
(2)TNF-αは、その受容体TNFR1に結合し、p38マイトジェン活性化プロテインキナーゼ(MAPK)経路の活性化を介して、テトロドトキシン(TTX)耐性Na+チャネルを増強する。
(3)IL-1bは、その受容体IL-1R受容体に結合し、p38MAPK経路の活性化を介して、テトロドトキシン(TTX)耐性Na+チャネルを増強する。
(4)IL-1bは、シクロオキシゲナーゼ-2系を介して、SPを放出する疼痛ニューロンを誘導することもできる。
[ET-1]
血管収縮作用を持つことが知られている21アミノ酸のペプチドである。
(1)ET-1を足底に皮下注射すると、ET-A受、受容体の活性化を通じて、疼痛行動を引き起こすことが示された。
※これらの知見を総合すると、上皮-神経細胞間疼痛経路の多様性が示される。
免疫細胞の役割:
慢性疼痛の原因となる免疫細胞には、肥満細胞(mast cell)、好中球、活性化マクロファージ、Tリンパ球など、さまざまな種類がある。ここでは、免疫細胞の概要と、免疫細胞が末梢感覚神経をどのように調節しているかを述べる。
[肥満細胞]
体内に広く分布し、解剖学的に感覚神経と共存していることが判明している。肥満細胞は、より多くの生理活性物質を含む多数の顆粒によって特徴づけられる。活性化すると、肥満細胞は、脱顆粒を起こし、細胞質顆粒の内容物を放出する。
これらの顆粒には、ヒスタミン、プロテアーゼ、セロトニン、顆粒球マクロファージコロニー刺激因子(GM-CFS)、IL-1,3,4,5,6、TNF-αなどのサイトカイン、CCケモカインリガンド(CCL)-1、CCL2、CCL3、CCL4などのケモカインが含まれる。
これらのメディエーターは、末梢求心性感作と炎症性疼痛につながる、感覚神経末端上の特異的受容体を標的とすることができる。
肥満細胞の脱顆粒は、ヒスタミン作動性経路と非ヒスタミン作動性経路を介して、侵害受容感覚も制御している。
[好中球]
組織障害に反応して、血液から組織に浸潤する、最も早く、リクルートされる自然免疫細胞である。
IL、プロスタグランディンE2(PGE2)、TNF-αなどの好中球から放出される炎症性メディエーターは、侵害受容ニューロンを感作することができるが、好中球と侵害受容性Never端末とのクロスストロークに関する知識は限られている。
末梢における好中球の蓄積を抑制することで、炎症後の疼痛を軽減できる。
[活性化マクロファージ]
そのクラスにより、痛みを調節することができ、その一つが、炎症性M1様表現型であり、もう一つが、抗炎症性M2表現型である。
ここでは、その炎症促進機能に着目する。
他のタイプの、免疫細胞を同様に、活性化マクロファージは、IL-1、NGF、TNF-αなど多くの炎症メディエーターを放出し、神経を感作する。
アンジオテンシンⅡ タイプ2受容体(AT2R)拮抗薬は、さまざまな疼痛モデルを持つ齧歯類の疼痛行動を軽減することが示されている。
傷害部位に存在するマクロファージではAT2Rが活性化されるが、DRGニューロンでは、活性化されず、活性酸素/窒素種の産生を誘発し、細胞損傷/疼痛感知イオンチャネルTRPA1の活性化を介して、侵害受容器を活性化する。
補体系成分C5aは、活性化マクロファージに依存して、NGFの放出を伴う、熱的および機械的痛覚過敏を誘発することができる。
NGFが侵害受容器の末端にある受容体に結合すると、侵害受容器上の、TRPV1受容体が感作され、痛覚過敏が誘発される。
貪食は、炎症の解消におけるマクロファージの重要な機能である。
ニューロプロテクチンD1(neuroprotectin D1: NPD1)による、マクロファージに発現する推定ペプチド性Gタンパク質共役型受容体(GPR)の活性化は、炎症性シグナル伝達から抗炎症性シグナル伝達への貪食と表現系の切り替えに関与している。
※免疫細胞が多くの慢性疼痛疾患において主要な役割を果たしていることを示唆するエビデンスが増えている。末梢に標的化した免疫細胞とそのメディエーターは、新しい治療の可能性を提供するかもしれない。
末端シュワン細胞の役割:
神経損傷後、シュワン細胞は損傷を迅速に検出して応答することができる。シュワン細胞は表現型の変化を受け、増殖し、軸索と相互作用して、ATP、成長因子(NGF、BDNF、ニューロトロフィン-3および4)、エリスロポエチン(Epo)、サイトカイン(TNF-α、IL-1およびIL-6)、ケモカイン(CCL1およびCOX2)などの多数のグリアメディエーターを産生する。
シュワン細胞によって放出されるTNFα、IL-1α、IL-1βなどの一部のグリアメディエーターは、マクロファージを動員することができ、これは間接的に軸索の興奮性の状態に影響を与える可能性がある。
ニューロンへのシグナル伝達に加えて、シュワン細胞によって放出された一部のメディエーターは、自己分泌的にシュワン細胞自体にも作用する。
例えば、NGFはシュワン細胞と軸索の両方を標的とすることがでる。この栄養因子は、DRGニューロンの軸索上の高親和性トロポミオシン関連キナーゼA受容体(TrkA)や、シュワン細胞上の低親和性神経成長因子受容体(LNGFR)に作用することができる。
神経損傷に応答して、シュワン細胞は細胞死を減少させ、軸索変性を防ぐシグナルを受け取り、それによって慢性的な痛みの状態を改善する。これらには、EpoによるEpo受容体(EpoR)、LDLによる低密度リポタンパク質(LDL)受容体関連タンパク質(LRP1)、NMDA受容体(NMDA-R)、およびグルタミン酸によるγ-アミノ酪酸B型(GABA-B)受容体の活性化が含まれる。
ATPによるP2X7受容体の活性化、損傷関連分子パターン(DAMP)によるToll様受容体2(TLR2)、LPAによるリゾホスファチジン酸1(LPA1)受容体]、およびシュワン細胞のシグナル伝達によるTRPA1(LPA1)受容体は、軸索変性を悪化させ、慢性疼痛を悪化させる可能性がある。
ErbBシグナル伝達は、リガンドの活性化に応じて、疼痛に対してプロペインまたはアンチペインになる。髄鞘形成シュワン細胞におけるニューレグリン/ErbBを介したErbBリン酸化シグナル伝達の欠損は、髄鞘形成低下と機械的刺激に対する過敏症を引き起こす。ハンセン病は、患者と動物モデルの両方に神経障害性疼痛を誘発する。M. lepraeは、ErbB2およびErk1/2に直接結合して活性化し、脱髄を誘導することが示された。この脱髄は、感覚の変化や慢性的な痛みにつながる可能性がある。
シュワン細胞の変性は、隣接する軸索にも影響を与える可能性がある。第 4 および第 5 腰椎腹側根 (L4/5 腹側根切開術) モデルの切断は、ウォーラー変性が有髄遠心性線維のみに制限され、無髄軸索を温存し、損傷していない C 線維を取り巻く Remak Schwann 細胞 (ただし、髄鞘形成シュワン細胞ではない) の有糸分裂を引き起こす。したがって、シュワン細胞の異常な活動は、細胞が直接被覆する軸索を超えて広範な影響を与える可能性がある。
侵害受容ニューロンは、グリア被覆のない皮膚の遊離神経終末として存在し、有害な刺激によって直接活性化されると長い間考えられてきた。しかし、「侵害受容性シュワン細胞」と呼ばれる特殊な皮膚グリア細胞タイプが最近特徴付けられ、表皮-真皮境界に侵害受容線維を持つ網目状の器官を形成した。これらの細胞の選択的活性化は、侵害受容ニューロンの活性化を引き起こし、痛みを引き起こした。侵害受容性シュワン細胞または侵害受容神経のアブレーションは、相手の収縮につながる可能性があり、機械的、冷たく、熱的な痛覚過敏を含む神経因性疼痛様過敏を引き起こす可能性がある。これらの末端グリア細胞は、軸索を包む細胞と同様に、疼痛シグナル伝達の開始と伝達において重要な役割を果たす。
全体として、侵害受容性感覚ニューロンは、DRGの末端枝、軸索、および細胞体の周囲でグリア細胞によって覆われています。これらのグリア細胞は、健康な痛みの状態と慢性的な痛みの両方の条件下で細胞外環境を調節する。これらの細胞の活性化は、無数のメディエーターの放出を介して侵害受容感覚を強化するために必要かつ十分である。
交感神経萌芽(sprouting)の役割:
通常、交感神経は真皮下部と皮下組織を神経支配し、真皮上部と表皮を占める侵害受容感覚ニューロンとは交差しない。
しかし、慢性疼痛では、異常な発芽交感神経が末梢の部位で疼痛感覚ニューロン終末とクロストークし、疼痛信号に影響を与える可能性がある。
感覚神経節と比較して、交感神経節における神経成長因子受容体TrkA(Ntrk1)の発現と濃縮が有意に増加した。
NGFとその高親和性TrkA受容体は、交感神経の発芽を導く上で重要な役割を果たす可能性がある。
交感神経軸索の側副発芽は、NGF の利用可能性に依存する。
神経損傷後のこれらの細胞タイプでNGF発現の増加が観察されていることから、NGFの潜在的な供給源には、マクロファージやシュワン細胞などの非ニューロン細胞が含まれる可能性がある。
臨床的には、星状神経節ブロック、腰椎交感神経ブロック、腹腔神経叢ブロック、上下腹部ブロック、および神経節インパー ブロックを使用して交感神経の活性化をブロックすると、複雑性局所疼痛症候群 (CRPS) 患者の約 50% で慢性疼痛を緩和できる。
皮膚アドレナリン作動性受容体/アドレナリン作動性過敏症が交感神経維持疼痛(SMP)のメカニズムに関与している可能性がある。
アドレナリン受容体は、α1-およびα2-アドレナリン受容体を含むα-アドレナリン受容体と、β1-、β2-およびβ3-アドレナリン受容体を含むβ-アドレナリン受容体からなるGタンパク質共役受容体のクラスである。
イソレクチンB4(IB4)+およびNF200+求心性線維に対するα1-アドレナリン受容体(α1-AR)の発現亢進は、末梢神経損傷後のラットの損傷した坐骨神経、真皮神経束、および真皮乳頭状で見られた。
複合性局所疼痛症候群(CRPS)に苦しむ患者の皮膚の神経線維で、α1-AR発現の上昇が観察された。
α2-アドレナリン受容体が発芽した交感神経と感覚終末との間の相互作用に関与している。α2-受容体の位置については議論中です。
これらの受容体は節後交感神経ニューロン(PGSN)の末端に位置している、他の研究では交感神経には存在しないことが示されています。
神経損傷後の末梢神経におけるβ-アドレナリン受容体の発現と位置も、包括的に報告されていない。
慢性疼痛に関しては、副交感神経系の発芽を調べた研究はごくわずかです。ラット無毛皮膚の感覚除神経後に真皮上部に発芽する副交感神経線維は、精神神経の切断後および精神神経の慢性狭窄損傷(CCI)]後に観察されました。慢性的なcyclpphosphanmide誘発性膀胱炎ラットモデルでは、CGRP+感覚神経と副交感神経の両方の発芽がラット膀胱の粘膜で観察された。
要約すると、自律神経系と感覚系は一般的に健康な条件下では重ならないが、交感神経はさまざまな慢性疼痛条件下で異所性に芽を出し、侵害受容神経終末を直接活性化/増強/感作できることがわかっている。これらの知見は、末梢性副交感神経の発芽が慢性疼痛の根底にある重要な経路である可能性を示唆している。
Zheng Q, Dong X, Green DP, et al: Peripheral mechanisms of chronic pain. Med. Rev. 2(3): 251-270, 2022
| ヒトの痛覚に関連する皮膚受容体ヒトの痛覚に関連する皮膚受容体 |
| ASIC3: |
| PAR2: |
| P2X3: |
| TRAAK: |
| TREK1/2: |
| TRPA1: |
| TRPM8: |
| TRPV1: |
| VR1: |
| リストはすべてを網羅するものではない。 |
Dubin AE, Patapoutian A. Nociceptors: the sensors of the pain pathway. J Clin Invest. 2010;120:3760–3772
Gold MS, Gebhart GF. Nociceptor sensitization in pain pathogenesis. Nat Med. 2010;16:1248–1257
Peppin JF, Albrecht PJ, Argoff C, et al: Skin Matters: A Review of Topical Treatments for Chronic Pain. Part One: Skin Physiology and Delivery Systems. Pain Ther 4: 17-32, 2015
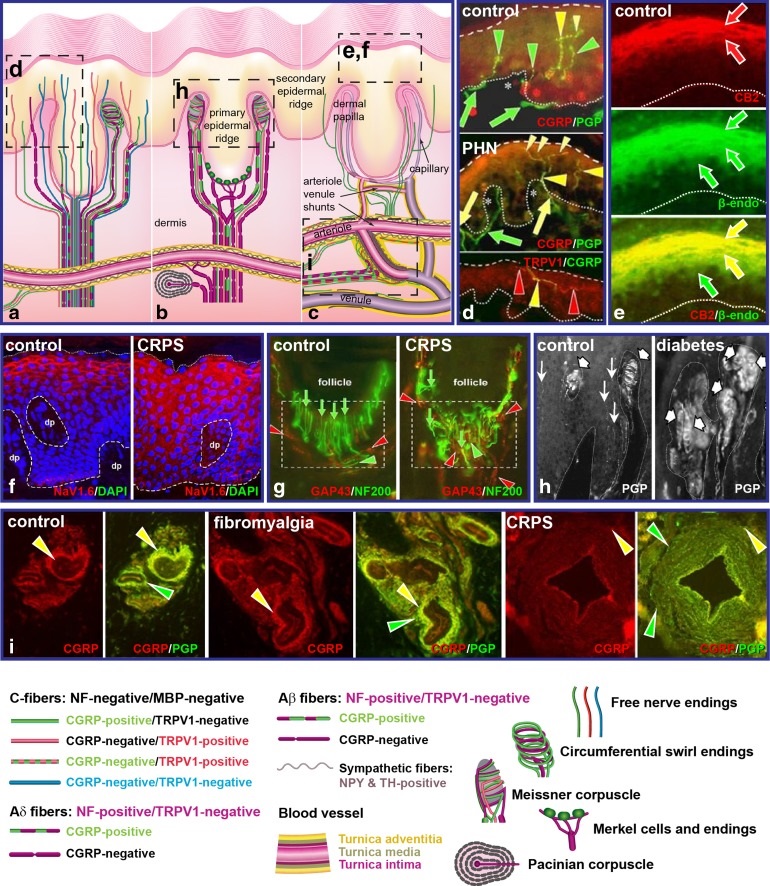
図 ヒトとアカゲザルの皮膚生検による多分子免疫蛍光法評価により検出された慢性疼痛関連病態の一例
図の説明
図のa-c:複数の研究結果をまとめたものに基づく、正常な無毛皮膚の指先部のすべての神経支配の模式図
図の e:角化細胞(ケラチノサイト)は、感覚伝達や、表皮の感覚終末における活動の調節に関与する痛み特性および鎮痛特性を有していることが示される。鎮痛機構には、上層部のケラチノサイトの間で、β-エンドルフィンの発現があり、これは、エンドセリン受容体B(ETB)とカンナビノイド2受容体(CB2)の活性化によって放出され、上層部の同じケラチノサイトで共発現する。
図の f:β-エンドルフィンの放出は、μ-オピオイド受容体、G-タンパク質活性化内向き整流カリウムチャネル(GIRK)、炎症性神経ペプチドであるカルシトニン遺伝子関連ペプチド(CGRP)およびサブスタンスP(SP)を発現する、表皮終末の活性を抑制する可能性がある。
アルギン酸のメカニズムには、ケラチノサイトのATP産生とCGRPのβ-アイソフォームが含まれ、これらはケラチノサイトに発現する、電位依存性ナトリウムチャネル(Nav)の活性化を通じて放出される。
ATPの放出は、プリン作動性受容体(例;P2X3)を活性化し、主にCGRPとSPを欠く表皮の末端に発現している可能性がある。
最近の研究では、帯状疱疹後神経痛(PHN)複合性局所疼痛症候群(CRPS)の患者の痛みを発する皮膚から得られたケラチノサイトの神経化学的性質は、痛み成分の過剰発現と、鎮痛成分の過小発現に偏っていると報告された。
図の b, g:個々のマイスナー小体や毛状複合体は、通常、複数のAβ線維の末端からなる複合体の高秩序な形態を持ち、数種類のC線維が混在していることが明らかになった。
さらに、Aβ線維とC線維の末端は、通常、侵害受容器に典型的に関連する免疫化学的に検出された特性を発現している。これらの特徴は、マイスナー小体と毛状複合体が、侵害受容器な関与をもつ可能性のある感覚統合の場であることを示している。
慢性疼痛のパラドックスの一つである、疼痛部位でも無感覚を示すという一般的な訴えは、Aβ線維の神経支配が失われている可能性が高いことに起因している。ししかし、2型糖尿病が自然に発症したアカゲザルと手足の切断を必要とした数名のCRPS患者を対象とした広範な皮膚サンプルの評価から、マイスナー小体と毛状複合体は存在したが、深く無秩序であった。
これらのデータから、しびれは、神経支配の消失ではなく、この神経支配による異常な活動によるものであることが示唆され、CRP患者で観察された、異常な毛状複合体は、毛のわずかな動きによる、深い機械的アロディニアに寄与している可能性がる。
図のc, i:CGRP/SPを含む求心性神経は、軸索反射を介した、真皮上部の毛細血管と毛細血管前細動脈の血管拡張と透過性の増加、および激しい熱に対する痛みの感覚に関与していることが示唆されている。しかし、真皮深部のおける細動脈や、細動脈―細静脈シャント(AVS)に対する広範で高密度の感覚神経支配に対する研究は少ない。
これらの深部血管の神経支配に対する、注目の多くは、最近、同定された、コリン作動性偶発事象を伴う、主に、ノルアドレナリン作動性の交感神経の神経支配に焦点が当てられている。
より深い結果の感覚神経支配は、いくつかのタイプのCおよびAδ線維からなり、そのほとんどが、CGRPおよびSPも発現している。この神経支配は、血管ダイナミクスのモニタリングの関与していると推定され、交感神経を介した血管収縮に対抗する、血管拡張エフェクターの役割を果たすと考えられる。
これらの細動脈やAVS求心性神経も、痛みを含む、意識的な触覚、知覚に寄与している可能性がある。皮膚血管系における感覚、交感神経支配の収束は、皮膚において、この神経支配の深い病態が観察される、CRPSや線維筋痛症などの慢性疼痛状態における、発汗障害や交感神経の関与の特異的な部位である可能性がある。
Peppin JF, Albrecht PJ, Argoff C, et al: Skin Matters: A Review of Topical Treatments for Chronic Pain. Part One: Skin Physiology and Delivery Systems. Pain Ther 4: 17-32, 2015
